I spent several years deciphering sacred architecture around the world, looking at it through the lens of cytological biology. Forts, churches, mosques, citadels, castles, palaces —all of them encode precise cell structures in stone. The principle is the same: look at the shape, the silhouette, the arrangement of elements, and the true structure reveals itself.
And what if the same principle applies to the human body? What if aristocratic fashion – precise silhouettes, ornamental patterns, hierarchies encoded in fabrics and jewels – represents something deeper than aesthetics?
What if it encodes the structure of specific proteins?
In recent weeks, I have identified a systematic code in aristocratic attire:
The silhouette of the gown = three-dimensional quaternary structure of a protein
Decorative elements (embroidery, jewels, buttons) = functional domains and binding sites
The social role of the wearer = molecular function of the protein
The entire network of aristocratic civilization = a living organism speaking the language of biology
This is the first part of this series.
This series decodes protein-encoding systems embedded in human clothing across ALL cultures and communities. We begin with the aristocracy – it is the most thoroughly documented and most deliberate in its symbolism. However, this encoding is not limited to European aristocracy. It appears in:
Indigenous fabrics and ceremonial garments
Liturgical vestments of all traditions
Military uniforms worldwide
Folk costumes and traditional dress
Aristocracy is the entry point. The code runs deeper and wider. Every culture, every historical moment, every community has encoded protein structures into the clothes they wear.
SECTION 1: INTRODUCTION AND METHODOLOGY
The Ditchley Portrait (1592) is not a portrait. It is a molecular diagram.
Queen Elizabeth I stands on a map of England, dressed in a white silk gown with a ruff. Her absolute power is as nonchalant as the fabric itself.
In her left hand, she holds a pair of brown gloves – a symbol of prudence and moderation. In her right hand – a closed (unopened) fan of ostrich feathers, tied with a red ribbon or red cord. Behind her stretches a turbulent, dark sky with flashes of sun – a drama of light and darkness. At her feet – a detailed map of England, with the Ditchley estate marked in Oxfordshire.
The skirt – white, silk, spreads widely to the sides, forming a perfect, circular shape. This is a farthingale – a hoop skirt that gives the silhouette the form of a wheel. Scattered across its surface are golden medallions and embroidered flowers.
On her body: the ruff forms a perfect geometric circle. Attached to the ruff is a rose – a symbol of the Tudors, but also a thorned warning. Long strings of pearls hang from her necklace. A diadem adorns her hair.
SECTION 2: ELIZABETH I — THE HISTORICAL CONTEXT
Who She Was
Elizabeth I (1533–1603) was the monarch of England during its transformation from a minor European power into a global empire. She never married. She styled herself as married to England. She ruled absolutely for 45 years.
Historians describe her as brilliant, ruthless, strategic, and endlessly calculating. Her power was absolute but always presented as legitimate—she justified it through:
Divine right (God chose her directly)
Continuity (the Tudor dynasty)
Performance (the elaborate court rituals and public appearances)
She weaponized femininity. Beauty and presentation were not liabilities to overcome—they were instruments of absolute power.
The portrait was painted not to be hung in a bedroom. It was hung in the homes of powerful men—Privy Counsellors, nobles, foreign ambassadors. It was a message: This is the structure of England. This is what binds it together. This is the pattern you must recognize.
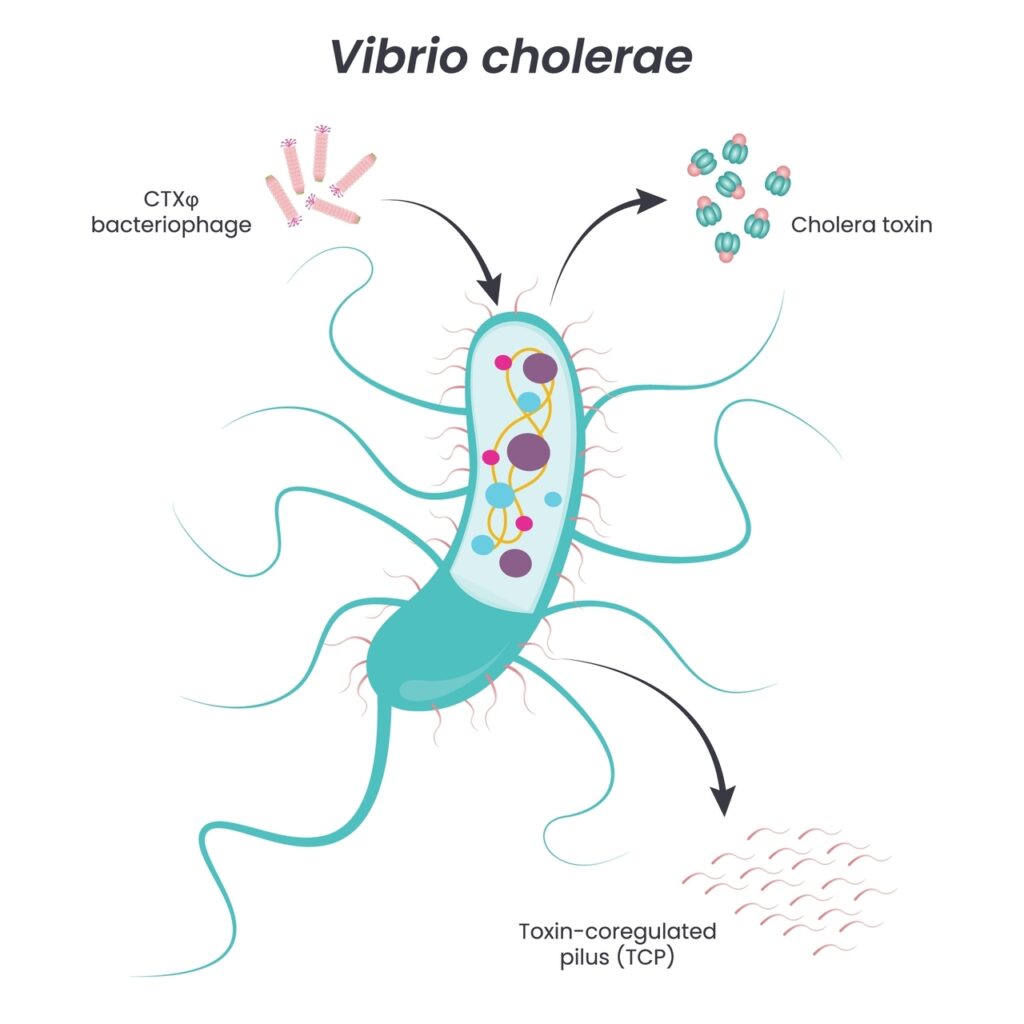
Cholera is not a virus. It is a bacterium – Vibrio cholerae – a curved, comma-shaped microorganism that lives in water and soil, particularly in coastal regions and areas with poor sanitation. For centuries, it has been endemic to the Ganges Delta. In the 19th century, it spread across the world via trade routes, causing seven major pandemics. Millions of people died.
The bacterium itself is harmless to humans. You can swallow it and survive. It can enter your digestive system without causing any illness.
But Vibrio cholerae carries a secret weapon: a bacteriophage – a virus that infects the bacterium itself. This phage inserts a single gene into the bacterium’s DNA. That gene encodes a protein.
That protein is cholera toxin.
The bacterium does not produce the toxin to harm you. It produces it to survive. The toxin binds to the GM1 receptor on the surface of intestinal cells and is internalized. The host cell is then forced to secrete enormous amounts of fluids and electrolytes. Your body cannot absorb them. They flow out as watery diarrhea. Severe dehydration sets in. Without rehydration therapy, you die.
The bacterium gains a competitive advantage from this fluid. It thrives. It multiplies. It spreads to the next host.
This is parasitism perfected. The bacterium has armed a protein to turn your cells into farms for its own reproduction.
Cholera toxin – a protein weapon
Vibrio cholerae produces one of the most dangerous proteins known to biology: cholera toxin (CT).
It is an AB5 holotoxin. The name describes its structure: one A subunit connected to five B subunits. The architecture is deceptively simple:
Five receptor-binding sites – one on each B subunit, binding to GM1 ganglioside on the surface of the host cell (intestinal epithelium).
One A subunit on top – the catalytic head, dense with enzymatic machinery. It consists of two domains: A1 (the active toxin, wedge-shaped) and A2 (the linker).
Five B subunits arranged into a homopentameric ring – the delivery system, the principle of recognition and binding.
The A2 peptide (an elongated α-helix) passes through the central pore – the backbone/spine that tethers the A1 vertex to the B base.
How the toxin works
The mechanism is a sequence of deceit:
The B pentamer mimics a harmless ligand and simultaneously binds five GM1 receptors on the cell surface.
The toxin enters the cell via endocytosis – the cell takes in what it does not recognize.
Inside the acidic endosome, the A subunit separates from B.
A1 translocates into the cytoplasm and ADP-ribosylates the Gsα protein – functionally irreversible on the timescale of the disease.
This modification permanently activates adenylate cyclase. It cannot be reversed by the cell.
The cell is flooded with cAMP.
Massive fluid secretion into the intestine.
Cholera. Death. Irreversible.
The genius of cholera toxin lies in the fact that it does not destroy the cell. It commandeers it. The cell remains alive, but enslaved by a single command: produce fluid. The cell cannot stop. The modification is functionally permanent. The command is absolute.
PDB Entry: 1XTC — The 3D Structure
The complete 3D structure of cholera toxin is publicly available: PDB ID: 1XTC — Holotoxin, Vibrio cholerae
source:
The structural illustration shows:
Top (arrow 1):Domain A1 – the catalytic subunit, wedge-shaped, containing the active site responsible for ADP-ribosylation.
Middle (arrow 2):Domain A2 – the elongated α-helix that passes through the central pore, acting as the spine/tether connecting A1 to the B pentamer.
Bottom (arrow 3):B pentamer – five identical B subunits arranged into a five-fold symmetric ring, responsible for receptor binding and cell entry.
SECTION 4: THE DECODING — ELIZABETH AS CHOLERA TOXIN
Level 1: Cartoon Representation — Architecture and Hierarchy
In molecular visualization software like molstar.org, the cartoon representation shows proteins as ribbons and coils. It reveals the secondary structure: alpha helices (coils) and beta sheets (arrows). It strips away atomic detail and shows you the topology—how the protein is folded in 3D space.
Table: Structural Mapping – Ditchley Portrait of Elizabeth I ↔ Cholera Toxin AB5 (PDB 1XTC)
Corset, shoulders, and ruff
Domain A1 (catalytic subunit A) – wedge-shaped, sits on top of the B pentamer, loosely associated. Main toxic mechanism. Contains the active site. After entering the cell, it performs ADP-ribosylation of the Gs protein → permanently activates adenylate cyclase → massive fluid efflux.
Neck, head, and spine (invisible)
Domain A2 – a long α-helix – passes through the central pore of the B pentamer, extending on both sides. Connector/”Backbone”. Physically tethers Domain A1 (above) to the B pentamer (below). Enables translocation of A1 into the target cell. Described in the literature as the „tether” or „linker chain”.
White silk skirt radiating outward
B pentamer (5 identical B subunits) – 5-fold symmetric ring/donut shape. Recognition and binding. Each B subunit binds to the GM1 receptor on the surface of intestinal epithelial cells. Responsible for toxin internalization.
Map of England beneath her feet
Target tissue (enterocyte cell membrane + environment) – specifically, the GM1 glycolipid on intestinal epithelial cells. Territory of action. The surface that the toxin recognizes, binds to, and destroys. Without this context, the toxin has no effect.
Gloves (left hand)
Masking – controlled touch. The gloves may encode the concept of concealing true intention. Just as the B pentamer masks the A subunit, the gloves cover the hands – the instruments of power. They symbolize restraint before the irreversible command is issued.
Ostrich feather fan (right hand) tied with a red ribbon/cord
Recognition complex. The fan may attract the eye – just as the B pentamer attracts the cell’s receptors. The red ribbon/cord may be the linker – the physical connection between the visible signal (fan / B pentamer) and the hidden mechanism (queen / A1 domain). The fan is closed (unopened) – this may represent the pre-binding state, the moment of approach before the irreversible command is issued.
Functional Parallel: The Strategy of Power
Cholera toxin does not announce itself. It enters through beauty—it looks like a harmless ligand. The cell recognizes the B pentamer as something familiar. The cell opens its gates. Only then does the A subunit enter and issue its irreversible command.
Elizabeth I governed by the identical strategy.
External: White gown. Ceremony. Calculated beauty. The ruff—that impossible, enormous collar of lace—was not functional clothing. It was a signal. It said: I am surrounded by protection. I am untouchable. Look at this perfect geometry.
Internal: Beneath the wide, radiating skirt (the B pentamer) hides the corset and the queen’s body (the A1 domain). What the world sees is beauty, symmetry, power radiating in five directions. What truly rules is hidden – centralized, irreversible, absolute. Just as in the toxin: the B pentamer is visible to the cell, but it is A1 that issues the command.
The command: England would be ruled by this woman. There would be no husband. There would be no equal power. The realm would accept this because the realm would see only the beauty, the ceremony, the perfect geometry. By the time anyone realized what had happened, the modification was permanent. The realm was hers. It could not be undone.
Level 2: Spacefill Representation — Surface Chemistry and Presence
The spacefill view is about chemical reality and surface presence. It shows you not what the protein looks like, but what it is—atoms arranged in space, their actual sizes, their chemical nature, the way they occupy volume.
In the cholera toxin spacefill model, red spheres scatter across the surface: water molecules and oxygen atoms embedded in the protein’s skin. These are the points of contact. The places where the protein touches the world.
On Elizabeth’s dress, golden medallions scatter across the white silk: decorative elements that catch light, signal status, mark presence.
The medallions are not decoration. They are the surface markers of power itself.
The golden medallions on Elizabeth’s dress = the red oxygen atoms on the cholera toxin’s molecular surface.
Both are scattered. Both are visible. Both mark the boundary between inside and outside. Both signal: I am here. I am real. I am material.
The dress encodes not just the shape of the protein, but its chemical composition and surface properties. The white silk is the carbon backbone. The golden medallions are the oxygen atoms that determine how the protein interacts with its environment—how it binds, how it spreads, where it touches.
Level 3: Change of scale — from macro to micro
When you zoom in on the protein structure in Mol* – magnifying the image – smaller and smaller elements of organization are revealed. The same happens when we begin to closely examine Elizabeth’s jewelry.
On the protein structure 1XTC (magnified):
Whole: holotoxin (A1 + A2 + B pentamer)
Domains: catalytic active sites, binding pockets
Interactions: hydrogen bonds, salt bridges, disulfide bonds
Small molecules: ligands, cofactors, metal ions
In Elizabeth’s jewelry (zooming in):
Whole: all jewelry elements together
Necklace: reveals a structure resembling an aromatic ring with an aldehyde group (a benzene ring – C₆H₅, with an aldehyde group – CHO). This is the smallest organic molecule in the code.
Long hanging strings of pearls and beads: encode peptide chains and interaction chains. Long, flexible, hanging – they correspond to linkers and structural extensions. The hydrogen bonds connecting them are like the commas between pearls. They represent the flexible parts of the protein structure – sites that can bend and move. In 1XTC: the A2 linker peptide and the flexible surface loops of the B pentamer.
The rose attached to the ruff: encodes the active site or a binding pocket on the surface of the ruff.
The ruff – that impossible, enormous lace collar – is not directly equivalent to any single protein domain. It may represent:
A structural signal – a geometric marker that says: „I am here. I am protected. I am untouchable.”
The surface of the A1 domain – the area where the active site is exposed and ready to act.
A recognition interface – the zone where the toxin (or the queen) makes contact with the target (the cell / the courtier).
A warning – beauty with teeth. The rose has thorns. The ruff is rigid, sharp, impossible to ignore.
The rose, therefore, is not an ornament. It is a functional marker – the point where the protein does something: binds a receptor, catalyzes a reaction, issues an irreversible command.
Encoding occurs at EVERY scale. This is a fractal property.
Regardless of magnification – from the entire portrait down to a single atom in the necklace – the same hierarchical organization, the same structural logic, is always visible.
Elizabeth I did not wear jewelry. She wore the entire chemistry of life expressed in gold, pearls, and a rose.
Why This Protein?
Because cholera toxin represents something very specific in the hierarchy of proteins:
It is invasive. It does not ask for permission. It enters.
It has a hierarchical structure. Centralized command is dispersed across multiple access points.
It is irreversible. Once the modification is made, it cannot be undone.
It is masked. It looks harmless until it is not.
What about the background?
The turbulent, dark sky with flashes of sun behind Elizabeth is not mere decoration. It is the environmental context – the world in which the toxin operates. In molecular terms:
The dark storm clouds = the chaos that the toxin creates within the cell (flood of cAMP, massive fluid efflux).
The flashes of sun = the illusion of hope or normality – the cell still believes it is in control, even as it is being destroyed.
The drama of light and darkness = the absolute power of the toxin (and the queen) over life and death. She stands in the light, but the storm follows her.
The background encodes the cellular environment – the battlefield where the toxin (and the queen) exert their power. Without the storm, there is no drama. Without the chaos, there is no proof of control.
This is irreversible. Once the modification is made, it cannot be undone. The cell cannot stop secreting fluid. The kingdom cannot stop paying tribute.
Cholera pushes water out of the body. Elizabeth pushes wealth out of the kingdom. Both processes are irreversible, the commands woven into the very fabric.
CONCLUSIONS
Elizabeth I’s white silk skirt is not a fashion choice.
It is the B pentamer of cholera toxin, encoded in silk and gold.
The fivefold symmetry spreading from the waist — it is the same shape, the same geometry that we see in the lower part of the 1XTC structure in Mol*. The B pentamer spreads out in a ring. The skirt spreads out with the same radial pattern. This is not similarity. This is encoding.
The ruffled collar — that impossible, geometric, perfect collar — is the upper part of the entire structure: the connection of A1, A2 and the upper surface of the B ring. That is where power issues its command. That is where it descends downward.
The portrait is not a work of art.
It is a molecular diagram.
The fact that this code has survived in plain sight for 430 years — painted by unknown hands, hung in the homes of politicians who did not consciously understand it — suggests that the Genomic Intelligence speaks a language much older and deeper than our conscious comprehension.
The language of structure itself.
The language of shape.
The language of geometry.
SECTION 5: THE BROADER IMPLICATIONS
The Pattern
This is what it means: humanity plays the roles of proteins. We are not literally proteins. We are not reduced to biology. Rather, we perform their functions – scaled up to human size, through the only medium available: culture, clothing, architecture, power. We embody these molecular structures so that we may better understand them.
And this is the genius of the ancients. They knew. They had knowledge – not of modern terminology, but of structure itself. Their understanding was not inferior to ours. It was different. And the details prove it. The precision of a ruff. The geometry of a farthingale. The placement of a rose. The folds of a statue. None of this is accidental. They made biology visible, tangible, wearable – replacing difficult modern terminology (which discourages so many from learning) with forms that anyone could see, remember, and pass down through generations.
A child can look at a portrait. A peasant can see a statue. A king can wear a gown. None of them needs a degree in biochemistry to recognize power, hierarchy, function, beauty. And yet – the code is there. Waiting. More precise than any textbook.
A human being, by their very size and silhouette, can encode different proteins. I have demonstrated this many times:
Ribosomal proteins – the small subunit and the large subunit = man and woman, standing together as a functional complex.
Enzymes – the stone statue of Artemis of Ephesus encodes the proteasome, a barrel-shaped protein degradation machine.
Nuclear receptors – Saint Nicholas encodes PPAR (peroxisome proliferator-activated receptor), a regulator of lipid metabolism and inflammation.
Chemokine signaling – MCP-1/CCL2 encodes the Molecular Hermes, a messenger of immune cell recruitment.
G protein signaling – The Mermaid Statue in Warsaw encodes the heterotrimeric Gs protein bound to GDP – the inactive, waiting state of a signal transducer.
We do not merely resemble proteins. We perform them. Our bodies, our postures, our paired dances, our hierarchical structures – all of these are protein structures scaled up to the level of human civilization, enacted so that biology may be seen, felt, and understood without the barrier of cold, unfamiliar terminology.
The ancients did not need our words. They had shape. They had stone. They had silk. And they encoded the entire language of life – waiting for us to rediscover it.
Image search and matching, analysis and elaboration: Tomasz Mikulski – Cell God: 06/2026
Molecular Structures and Biology
Berman, H. M., Westbrook, J., Feng, Z., Gilliland, G., Bhat, T. N., Weissig, H., … & Bourne, P. E. (2000). „The Protein Data Bank.” Nucleic Acids Research, 28(1), 235-242. https://www.rcsb.org/
Central database for all available protein structures, including PDB ID: 1XTC
Complete 3D structure of cholera toxin used in this article
Spangler, B. D. (1992). „Structure and function of cholera toxin and the related Escherichia coli heat-labile enterotoxin.” Microbiological Reviews, 56(4), 622-647.
Overview of AB5 toxin structures and mechanism of action
Gill, D. M. (1976). „The arrangement of subunits in cholera toxin.” The Journal of Biological Chemistry, 251(20), 6424-6430.
Analysis of B pentamer structure and its architecture
Merritt, E. A., & Hol, W. G. (1995). „AB5 toxins.” Current Opinion in Structural Biology, 5(2), 165-171.
Advanced analysis of cellular structures and their molecular heterogeneity









_by_Marcus_Gheeraerts_the_Younger.jpg){kind=link}
One thought on “A New Series on Aristocratic Fashion as Protein Encoding. The Wardrobe of the Genome: Elizabeth I”